Enjoy DatePsychology? Consider subscribing at Patreon to support the project.
The Growing Rift Between the Red Pill and Evolutionary Psychology
A recent paper published in Evolutionary Human Sciences documented how although manosphere and red pill communities claim to rely heavily on evolutionary psychology, they often misunderstand the content — and even misunderstand basic scientific concepts like what a hypothesis is (Bachaud & Johns, 2023). As a result, a handful of provocative hypotheses (some with relatively little support from the beginning) have filtered into manosphere ideology as if they were “facts” or “laws” about human sexual behaviors (assertions that evolutionary psychologists have not made regarding these hypotheses).
Bachaud and Johns (2023) reference the introduction to the Red Pill subreddit: “A large portion of Red Pill discussion revolves around evolutionary psychology.” It would be more accurate to say that a large portion of red pill discussion revolves around a caricature of evolutionary psychology. There is virtually zero overlap between the red pill and evolutionary psychologists. This is a truly remarkable feat for a community that claims to revolve around evolutionary psychology. As I have noted in past articles, the red pill subculture has produced no evolutionary scientists, has made no discoveries, and has contributed nothing to research within human behavioral fields.
Writing for the Boston Globe (see: How the incels warped my research), evolutionary psychologist Daniel Conroy-Beam recently described how the “scientific black pill” Incel Wiki cited his papers, but made assertions about findings that were not found anywhere in the results: “I thought, “That’s odd, I don’t remember ever publishing on dominance preferences — do the incels know my work better than I do?” No. I double-checked: That study didn’t even mention dominance preferences. Curiously overlooked in this whole wiki section on women’s preferences is the fact that kindness is repeatedly found to be among the most desired qualities in large-scale, cross-cultural studies of mate preferences.”
The dual mating strategy hypothesis is referenced ad nauseam in manosphere and red pill discourse and informs much speculation about “alphas” and “betas.” However, as Conroy-Beam explained, this hypothesis fell off almost a decade ago amidst the replication crisis:
“Part of the problem is that the dual mating strategy hypothesis was indeed a popular idea among evolutionary psychologists until about 2016. After that, it became one of the more prominent epicenters of psychology’s replication crisis, which revealed that large swaths of psychology research were based on unreliable findings. But even before this major setback, the dual mating strategy hypothesis was critiqued by some evolutionary psychologists like my friend and colleague Jim Roney. Nonetheless, Jim’s work gets hardly any play in manosphere circles, and the hypothesis has since morphed into a version quite unlike the one promoted by incels.”
In an interview with Slate back in 2018, Steven Gangestad — the researcher who came up with the dual mating hypothesis — also went on to say this about the dual mating hypothesis:
“The implications of this newer research have not escaped the founding fathers of the field. “In terms of overall effects, I don’t think there is anything,” admitted Gangestad in a recent interview, referring to his theory of women’s so-called dual sexuality. There may still be some difference in fertile-phase effects for women who are single and those with long-term partners, he said, “but the probability I would put on that is pretty low.” In other words, the man who literally co-wrote the book on human oestrus now believes that he and others in the field were the victims of pervasive problems in the way that the psychology was done—a bum steer that affected not just their research, but many others’ too. “When we wrote the book, we were drawing on a broad literature,” Gangestad told me, “but some of what we wrote was just garbage because we trusted all that work, including our own.””
Manosphere influencers are well aware of this, but stubbornly cling to old and non-replicable research nonetheless. The podcaster Rollo Tomassi has commented on my Twitter/X posts regarding the topic numerous times and referenced me by name in his shows on this topic, so I know that he knows better. In a recent video entitled “Why This IMPACTS Who Women Are ATTRACTED To!” he handwaves away the non-replication of the dual mating hypothesis and ovulatory shift research by claiming, in effect, that Steven Gangestad is afraid of the Department of Homeland Security and that feminists are responsible for failed replications.
Rollo Tomassi seems to be a big fan of the evolutionary psychologist Martie Haselton, another prominent researcher of ovulatory shifts in mate preferences, and name-drops her frequently on his shows. However, in her most recent review of ovulatory shifts, penned alongside Gangestad, Dinh, and Lesko (2022) for a chapter in the Oxford Handbook of Human Mating, here is what she wrote following two meta-analyses: one that found significant results (Gildersleeve et al., 2014) one that found null results for ovulatory shift preferences (Wood et al.). Both of these were back in 2014 and subsequently new replication attempts have been made:
“In light of contrasting results, a number of research teams have sought to assess the replicability of key findings in reasonably high-powered studies. Results have been mixed, though findings have been more negative than positive … What, then, can be concluded at this point in time? In our view, uniform null hypotheses, across all preferences, are unlikely … At the same time, claims based on early studies were likely too bold. If preference shifts exist, they are likely to be restricted to certain features. And even then, simple two-way interactions are unlikely to be large across women in general.”
Also from Gangestad, Dinh, Lesko, and Haselton (2022), the Dual Sexuality Framework (shifts associated with cycle phases) does not necessarily imply a dual mating strategy and there is good reason to believe that it servers a different purpose altogether — reinforcing pair-bonding:
“To evolve and be maintained, women’s extended sexuality must also afford special benefits. Paternity confusion, however, is an unlikely function. As pair-bonding and biparental care purportedly evolved in humans, females should have evolved to generally assure paternity of primary partners (e.g., Gavrilets, 2012). Symons (1979) and Alexander & Noonan (1979) proposed alternative functions: that women’s extended sexuality evolved (at least in part) to enhance pair-bonding and bolster male investment in offspring.”
Rollo Tomassi also seems to be unaware that ovulatory shifts in behavior broadly, ovulatory shifts in mate preferences, and the dual mating strategy hypothesis are not the same thing. For example, an increase in libido or sexual desire during ovulation (a robust and well-replicated effect) is not the same as a change in partner preferences (for example, a desire for more facially masculine men when ovulating, which does not replicate; see Gangestad, Dinh, Lesko & Haselton, 2022). Similarly, the dual mating strategy hypothesis — the hypothesis that infidelity is an adaptation to secure “good genes” from an extra-pair sire — is not synonymous with ovulatory shifts in partner preferences (for a comparison see the introduction of Thomas et al., 2021). If women are more intrasexually competitive during ovulation or shift their clothing choices (e.g. Zhuang et al., 2014) this is not evidence for a shift in mate preferences nor for the dual mating strategy hypothesis that infidelity evolved to secure “good genes.”
In this, Rollo Tomassi uses the same kind of “Russian nesting doll of misrepresentation” described by Daniel Conroy-Beam for the Boston Globe. In one Twitter/X reply to me (here) Rollo Tomasi posted a paper (Schleifenbaum et al., 2021) that found women feel more attractive before ovulation. The results do not show a shift in mate preferences during ovulation, nor do they touch on the dual mating strategy hypothesis in any way. The authors of the paper note that the results are consistent with the motivational priorities shift hypothesis — an alternative to the ovulatory shift hypothesis.
In a second example (here), Rollo Tomassi references a paper showing a shift in food preferences during ovulation (Nijboer et al., 2024). This may very well be a real effect. However, this does not support shifts in mate preferences during ovulation nor that infidelity is an adaptation to secure “good genes” during ovulation. He seems to believe that ovulation having any effect whatsoever upon human psychology is support for the ovulatory shift hypothesis or the dual mating strategy hypothesis, but it isn’t, nor do the psychologists claim it is within the research he cites. Alternatively, he might just expect that you will not read the papers and find out that he is misrepresenting their content.
A final word could be said about effect sizes. Effect sizes estimate the strength of a relationship between variables, or the size of a difference between groups. Influencers within the manosphere seem allergic to discussing effect sizes, or they don’t know what effect sizes are and how to interpret them. I don’t know which of the two. Effects for single variables in psychology tend to be small. If someone says “ovulation shifts preferences for muscularity in men” your immediate question should be “by how much.” Many effects are real, but are so small they are not meaningful by themselves. Further, research reporting small effects is less likely to replicate (Boyce et al., 2023).
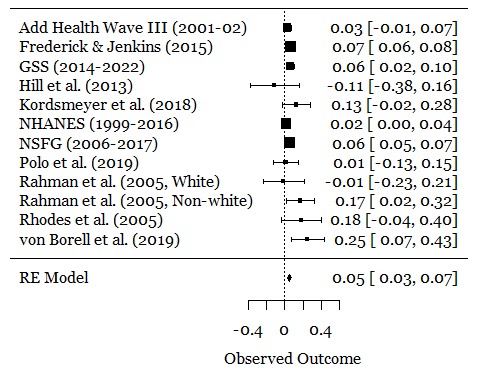
Setting aside all of the post-2016 failed replications of ovulatory shift research, we can go back to the original meta-analysis by Gildersleeve et al. (2014). Below are effect sizes for shifts in mate preferences:
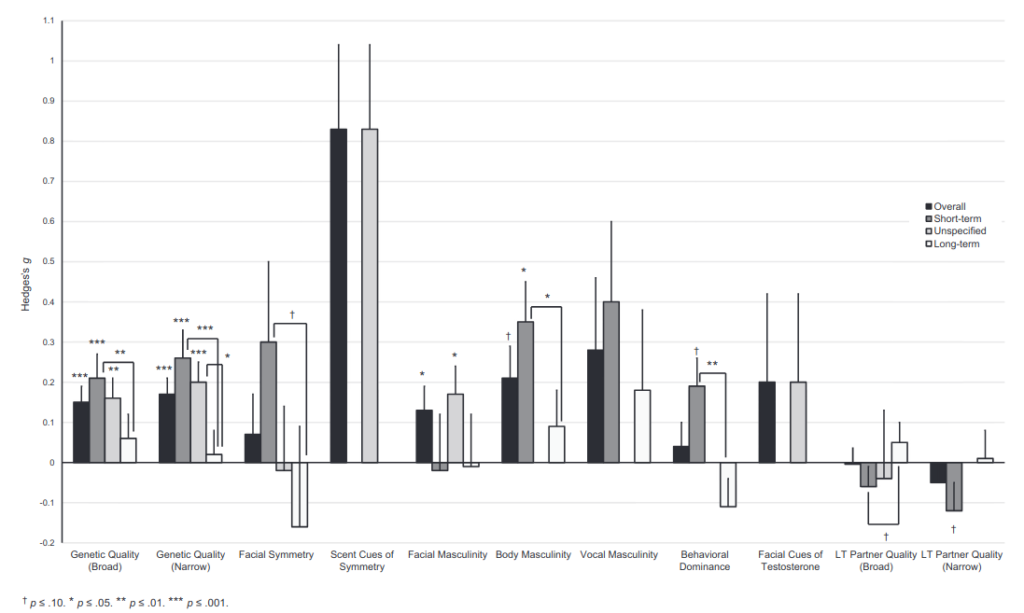
Hedges’ g for an ovulatory shift in female preferences of ancestral genetic quality in males across all relationship contexts was .15 and in short-term contexts specifically was .21. Real or not, these are quite small to build an entire ideology of “alphas and betas” around. Below is a visualization from the R Psychologist app to help you understand what the difference looks like:
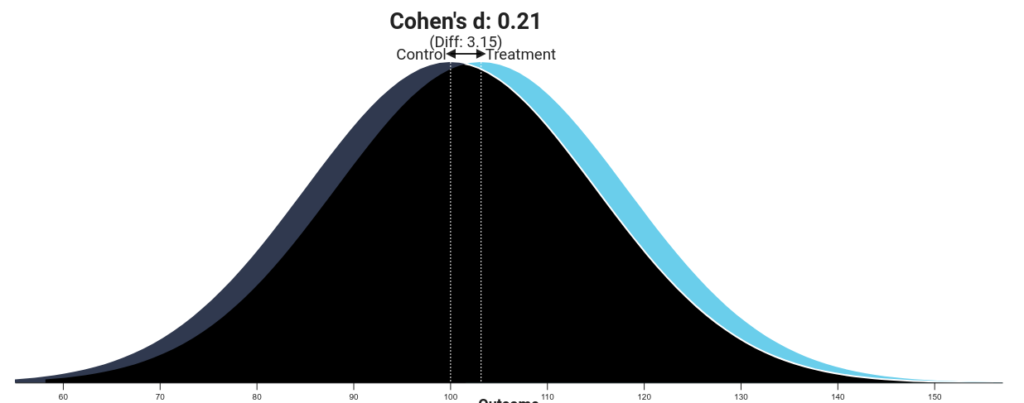
Gangestad, Dinh, Lesko, and Haselton (2014) never claimed that these were large effects — they report them as small in the meta-analysis — and you can see that mate preferences overwhelmingly overlap regardless of fertility status. The kind of man an individual woman desires is mostly the same regardless of where she is in her ovulatory cycle.
Steve Stewart-Williams covered a recent paper by Michal and Shaw (2024) on how laypeople interpret scientific findings when effect sizes are not reported (see: Effect Size Matters). Unsurprisingly, when effect sizes were not reported, participants tended to assume they were large (image below). Further, individuals low in numerical or math skills were more likely to endorse or act on small effects even after being told they are small.
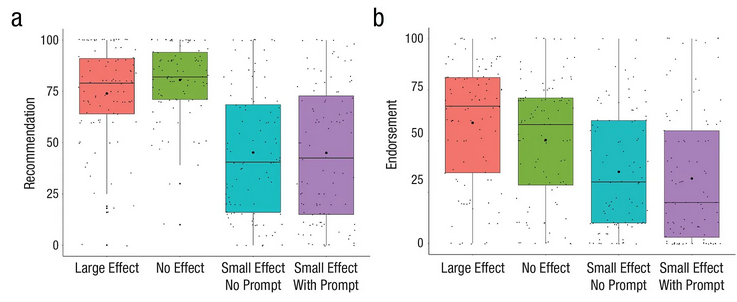
I have little doubt that numerous manospherians have formed the impression that, when listening to debates over ovulatory shifts or other topics in psychology, we are talking about large effects. In most cases we are not: the average effect size in psychology is .30. Research has never shown that women go from wanting a wimpy milquetoast “beta” during nonproliferative stages of the ovulatory cycle to an entirely different type of man when fertile. Mate preference shifts, if they are real at all, are small and subtle.
Monogamous Men: Are They “Betas?”
The title of this article was inspired by Rollo Tomassi’s red pill blog post “The New Polyandry” with a subheading of “Monogamy is beta.” In this a few claims are made:
- The “alpha” is attractive and is a fast life history strategist.
- Most men would seek unlimited sexual access to women if they could.
- The “beta” is unattractive and his strategy is monogamy; he is a slow life history strategist.
- Monogamy is a social norm from the Agrarian revolution, not an evolved behavior.
Evolutionary psychologists do not use the terms “alpha” and “beta,” but fortunately do have something that approximates the popular dichotomy: high and low mate value. We can also look at other behaviors that the manosphere and red pill subcultures group in with “alpha” and “beta” caricatures, such as parental investment. We can also examine if monogamy and parental investment are associated with status, attractiveness, and more.
Human beings have both physical and behavioral traits that are robustly desirable across cultures (Buss, 1989; Shackelford et al., 2005; Walter et al., 2020). Having more desirable traits results in higher mate value and corresponds to a given individual being more desirable to more members of the opposite sex, or even the same sex (as desirable traits also tend to be consistent across sexual orientation) (Stephen & Luoto, 2021).
The magnitude placed on specific traits varies by mating context. For example, there is a premium on physical attractiveness in short-term (casual sex) mating contexts (Gangestad et al., 1993; Buss, 2023). However, people often make the mistake of believing that this means some traits are desirable in short-term contexts that are undesirable in long-term contexts. The same traits that are desirable in short-term contexts are also desirable in long-term contexts and vice versa (Buss, 1989; Buss, 2023; Li & Choy, 2023). In fact, short and long-term mate preferences are highly correlated. For example, physical attractiveness is still highly desirable in a long-term mate, even if it is weighted slightly higher in short-term mating contexts. There seem to be few, if any, traits desirable in one mating context that are undesirable in the other.
Further, although evolutionary psychology has often framed mating as short or long-term (for example in Sexual Strategies Theory, see Buss, 2023), much of human “short-term” mating may actually fall into an “intermediate” category with varying levels of long-term commitment (Jonason & March, 2023). For example, “booty calls” and “friends with benefits” relationships are ongoing but have lower expectations for exclusivity and commitment. As a result, we should expect traits conventionally associated with long-term mating preferences to also be valuable in what is often described as short-term mate selection. This is precisely what we see: traits that facilitate long-term relationship formation such as competence, compassion, compatibility, kindness, altruism are desired even within short-term mating contexts (Jonason & March, 2023).
Sociosexuality and Fast/Slow Life Histories
There are individual differences in the traits that constitute mate value. Further, there are individual differences in orientation toward short and long-term mating (Simpson & Gangestad, 1991; Buss & Greiling, 1999). Long or short-term mating orientation may correlate with what is called life history strategy. Individuals with a slow life history strategy have a stronger orientation toward long-term mating (Figueredo et al, 2004; 2006). A slow life history strategy also involves higher investment in offspring as well as in individual quality (Figueredo et al., 2006) — these individuals invest more time into themselves and into cultivating high mate value within themselves (physical and behavioral attractiveness). Conversely, individuals with a fast life history strategy, and/or a higher short-term mating orientation, invest more effort into mating (Figueredo et al., 2004).
This is the broad theoretical difference between fast and slow life history strategies: slow life history strategies dedicate more time toward somatic effort, or individual survival, while fast life history strategies dedicate more time toward mating effort, or reproduction. Both represent alternative routes to reproductive fitness. For slow life histories, increased longevity, individual health, choosiness of mates, and high parental investment facilitates reproductive fitness. For fast life histories, eschewing mate-choice selectivity and parental investment allows individual organisms to reproduce more. This is uncontroversial when contrasting animal species (MacArthur & Wilson, 2001). Human beings are a slow life history species (few offspring, long gestation periods, high investment in offspring) when contrasted with rats (many offspring, low gestation period, low investment in offspring). More recently, life history theory has been applied to individual differences within species (e.g. humans).
The premise of life history theory applied to individuals and its relationship with sociosexual behavior proposes that short-term mating in human beings increases offspring quantity (Ellis et al., 2009). After all, the more sexual partners a woman has the more offspring she will conceive, right? Maybe not, according to Dinh, Haselton, and Gangestad (2022; see also Dinh & Gangestad, 2024):
- Fertility rates among foragers are determined by interbirth intervals (in effect, how closely back-to-back can a woman give birth). Intervals are lowest when there is high parental nutritional provisioning: “Females can more effectively increase offspring quantity by forming stable long-term partnerships.”
- “The assumption that female willingness to engage in uncommitted sex facilitates high offspring quantity or genetic diversity is biologically infeasible.” Dinh et al. (2022) explain that sex with multiple partners doesn’t produce multiple offspring in most cases. Further, it does not produce more offspring than sex with a single pair-bonded partner and would be counterproductive toward genetic diversification. On top of that, it undermines parental certainty and risks the loss of parental provisioning, which subsequently leads to lower offspring quantity.
In a recent review of life history theory, Dinh and Gangestad (2024) note that there is little theoretical or empirical reason to believe that unrestricted sociosexuality promotes higher mate quantity or quality in hunter-gatherer/forager populations (in evolutionary psychology and anthropology, these populations are used to approximate the ancestral environment in which our mating psychology evolved). In other words, long-term mating results in having more offspring and, most likely, higher quality offspring. They conclude:
“In our view, these models have not offered compelling arguments for how proclivities to form concurrent or short-term sexual partnerships, as opposed to forming committed pair-bonds in service of biparental care, function to promote current versus future reproduction or offspring quantity versus quality in human ancestral environments. On the contrary, data across human foraging groups reveal that male provisioning speeds up women’s fertility rate and increases offspring quantity.”
Given this, it is important to note that a fast life history is not necessarily synonymous with an unrestricted sociosexuality or short-term mating orientation going forward. It is also not necessarily a more effective mating strategy. That said, we should not throw the baby out with the bathwater. Marco Del Giudice (2024) proposed an extended model of life history that rectifies some of the issues raised by Dinh & Gangestad (2024):

Many traits associated with fast and slow life history strategies are robust. In particular, associations between fast life history strategies and antisocial traits, as well as traits associated with lower desirability as a mate, are robust. We can still examine relationships between life history strategy, short/long term mating orientation, and mate value.
Mate Value & Life History Stratregy: Bivariate Correlations
First, a few studies assessing mate value and life history strategy directly:
Dillon et al. (2013) found positive correlations between life history strategy and mate value at .49 for the individual and .32 for the mate value of their romantic partner. Kennison et al. (2017) found life history strategy and mate value correlated at .31. Buunk and Massar (2021) found a correlation between life history strategy and mate value of .55. In McDowell and Starratt (2021) this relationship was .34 for men. Further, estimated heritability for the relationship between life history strategy and mate value was around 50% for most traits: being physically strong, physically attractive, dominant, trustworthy, and more. A slow life history strategy — commitment to long-term, monogamous relationships with high parental investment — was positively associated with mate value in all of these.
Mate Value & Sexual Partner Count
It may be counterintuitive that individuals who have more sexual partners could also have lower mate value. A minimum threshold of mate value must be met in order to find a sexual partner. Individuals of higher mate value are also more able to meet their mating goals. However, it is not the case that a universal mating goal for all men is simply maximizing one’s number of lifetime sexual partners. Individual differences in sociosexuality, disgust sensitivity, sensation-seeking, and a long list of additional personality traits shape the extent to which any given individual will desire sexual variety over a committed relationship. From life history theory, we also expect slow life strategists to produce offspring of higher somatic quality (for example height) (Del Giudice, 2024) and to be of higher somatic quality themselves, yet also expend lower mating effort, invest more in long-term relationships and parental care, and pursue fewer short-term sexual opportunities.
The important takeaway here — these traits correlate or interact with mate value, such that we are able to observe how men who are high or low in mate value behave on average. We tend to see many traits that are desirable in a male partner, both physical and behavioral, that are associated with both long-term and slow-life history strategies rather than exclusively with a short-term orientation.
A good illustration of how men who are low in mate value may nonetheless accumulate more sexual partners is found in the Dark Triad (see: Is the Dark Triad Really Attractive? A Review of the Literature). High DT individuals — who on average score low across a wide selection of mate value metrics, including facial attractiveness in some research — nonetheless accumulate more sexual partners. Why and how?
Men high in the Dark Triad have a stronger orientation toward forming short-term rather than long-term relationships, so they don’t spend extended periods of time in monogamous relationships (and are also less able to voluntarily maintain monogamous relationships; they are more likely to leave relationships, more likely to be broken up with, and more likely to be cheated on). They have lower disgust sensitivity and lower mating standards. As a result, men higher in the Dark Triad are less picky about who they have sex with and they choose less attractive mates. They also exert substantially more effort in pursuit of short-term mating opportunities (mating effort), as well as engage in more self-promoting and image-enhancing activities. The traits that constitute the Dark Triad are associated with a fast life history strategy.
Given that physical attractiveness is consistently among the top predictors of mate choice, both for short and long-term relationships, we might expect to see a strong relationship between physical attractiveness and number of lifetime sexual partners. However, this would only occur if most men were engaging in a similar mating strategy designed to maximize sexual partner count. In other words, if all that men really wanted was to seek out sexual novelty and maximize the number of women they have sex with, we should see a strong linear correlation between physical attractiveness and lifetime sexual partner count. Instead, here is what we see:
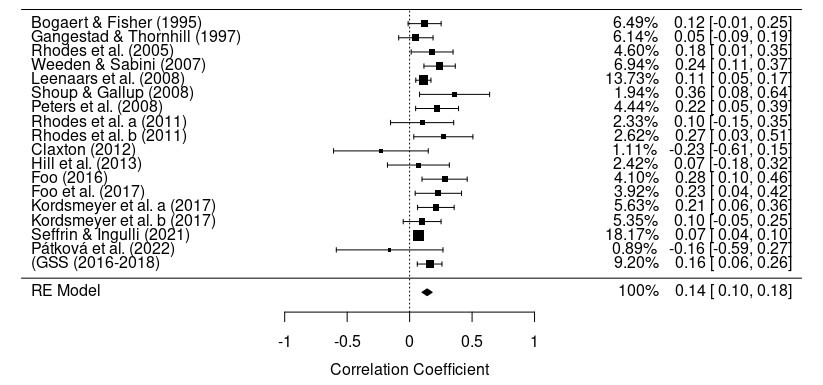
Physical attractiveness has only a small relationship with lifetime sexual partner count. This is what you might expect if most people simply enter and stay in long-term relationships. A man who is 9/10 in physical attractiveness should end up with many more sexual partners than a man who is a 4/10 — but only assuming both pursue short-term mating. We should expect to see a much stronger relationship between physical attractiveness and sexual partner count than a mere g of .14 if short-term mating were the norm for attractive men.
Here we see the same relationship in a meta-analysis of male height and lifetime sexual partner count (from @nuance_enjoyer on X/Twitter). This relationship is even smaller — it is practically nothing. This is what we would expect if taller men are following the same pattern as shorter men: serial monogamy and long-term relationship formation. This is a well-observed phenomenon in men; taller men are more likely to be married (Murasko, 2020; Steckel, 2009).
Height also shows a curvilinear relationship with reproductive success (Stulp & Barrett. 2016): men of average height are more reproductively successful than short or tall men. Height is positively selected for by women (at least in northern hemispheres, per Stulp & Barrett), so why don’t taller men have higher reproductive success when taller men are both preferred by women and have higher status on average? This reflects the life history trade-off between somatic and mating effort. Taller men are more likely to be slow life history strategists.
A recent meta-analysis of sociosexuality and male self-reported mate value also found only 6% of the variance in sociosexuality was explained by mate value (Arnocky et al., 2021). Men with a higher short-term orientation perceive themselves as having higher mate value, but as the authors explain this relationship is small enough that it could be entirely confounded by traits like narcissism. Importantly, mate value explains very little of individual differences in sociosexuality. Of all the things that make men more likely to pursue short-term mating strategies, having a high mate value is at best a tiny part of the picture.
In other words, not only are prime physical predictors of mate value (physical attractiveness and height) weakly or unrelated to sexual partner count, the desire to engage in a short-term mating strategy is also weakly related to mate value.
Mate Value and Infidelity
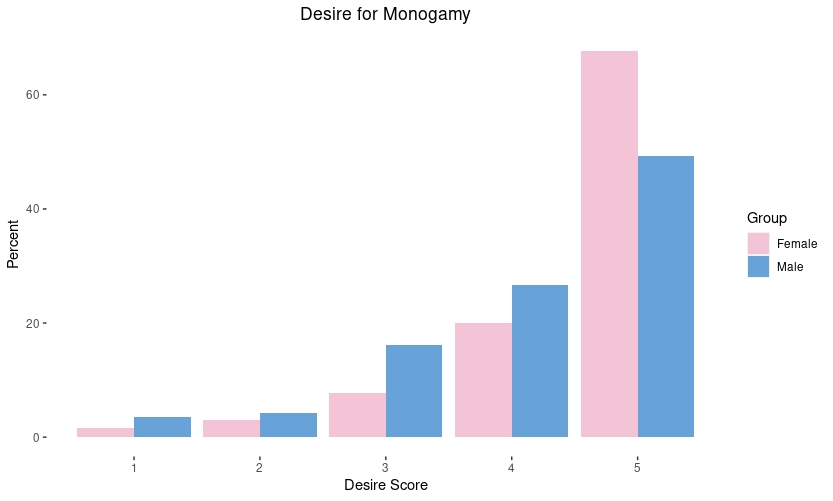
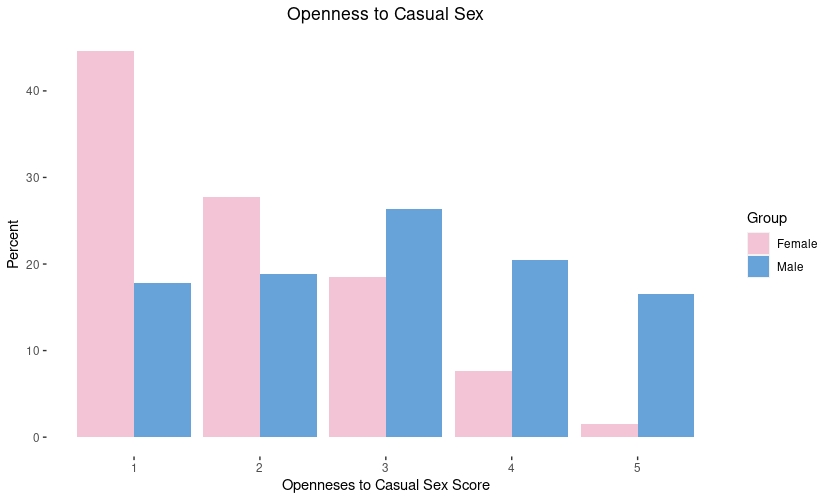
A common belief shared in the manosphere and red pill communities is that men would pursue short-term mating strategies if they could and thus men “settle” in long-term mating strategies because they cannot effectively pursue short-term relationships. In other words, women are naturally monogamous and men are naturally promiscuous. Not so, says evolutionary psychologist Steve-Stewart Williams: “Both sexes have the capacity for both long-term and short-term relationships, and both are about equally interested in long-term ones.” (See: 12 Things Everyone Should Know About Evolutionary Psychology). There is a large sex difference in sociosexuality with men being more unrestricted, but this does not mean men have a low interest in forming monogamous relationships (see: We Were Patriarchs: Male Mating Strategies and Intrasexual Competition). This is a pattern I am able to replicate in my own surveys as well:
Relatedly, a common belief shared in the manosphere is that men of high mate value are more likely to engage in infidelity. Research also does not support this claim or is, at best, mixed and conditional. Starratt et al. (2017) found a negative relationship between overall mate value and intentions to commit infidelity. Further, partner mate value (also associated with self mate value) was inversely associated with intentions to commit infidelity. This is important, because partner mate value is predictive of self mate value. In another recent study, Nascimento et al. (2024) found that success on online dating applications was not associated with infidelity. Further, the perceived availability of potential mates — the belief that you have a lot of people who would potentially date you — was not associated with infidelity.
This is not to say that mate value is unrelated to infidelity in all cases. High(er) mate value may predict infidelity when there is a mate value discrepancy, or a partnership between one individual of higher mate value and one individual of lower mate value relative to them (Buss & Shackelford, 1997; Nascimento et al., 2024). Further, mate value also predicts differences in mate retention behaviors (evolved strategies to thwart infidelity or the defection of a romantic partner) (Miner et al., 2009; Sela et al., 2017). However, this is not a statement about absolute or high mate value. Individuals of high or low absolute mate value may both have a gap between themselves and the mate value of a romantic partner. Because human beings mate assortatively for all traits associated with mate value (Conroy-Beam et al., 2019a, 2019b; Conroy-Beam, 2018), and because individuals of high mate value are able to more successfully meet their mating goals (selecting partners with more valuable traits), we should not expect more mate value discrepancies for individuals of higher mate value in relationships than for lower mate value individuals.
Individuals of higher mate value are also more likely to successfully mate poach — or steal a romantic partner away from another relationship (Schmitt & Buss, 2011; Arnocky, 2020). However, the key word here is successfully. Mate value did not predict unsuccessful poaching attempts. Individuals high in mate value may be more successful at poaching a mate, but are not necessarily more likely to attempt mate poaching. This could lead to the perception that high mate value individuals engage in unfaithful behavior more frequently simply because successful mate poaching attempts are more salient. Intrasexual competitiveness — which can be induced by low mate value and which is associated with fast life history strategies — predicted mate poaching attempts. In other words, it isn’t clear that more desirable mates are more likely to attempt affairs, but they are more likely to be successful when they try. Here mate value discrepancies also matter: the partners of high value individuals are less likely to be poached and individuals in relationships with a high value partner are less likely to engage in mate poaching themselves (Davies & Shackelford, 2017).
The evolutionary psychologist David Schmitt led a team of 121 psychologists across 53 nations to study mate poaching (Schmitt et al., 2004). In their seminal paper they described what kind of person mate poaches. To paraphrase:
Mate poachers are disagreeable, unconscientious, unfaithful, and erotophilic. They are lower in empathy. In the case of successful mate poachers, they are more open to experience, perceive themselves as more attractive, and have an unrestricted sociosexuality. Mate poachers also have a predisposition for narcissism and unfaithfulness that reflects a genetic orientation toward short-term mating.
Individuals who are the target of mate poaching, and even more individuals who have been successfully mate poached, share a remarkable similarity across the above traits. Here we see assortative mating once again at work: individuals attempt to select partners similar to themselves (even when those partners are already in existing relationships).
Most of the traits above are associated with low, rather than high, mate value.
Mate Value and Parental Investment
Robert Trivers classically observed across animal species that the lower-investing sex becomes less sexually selective and can increase their reproductive fitness by devoting more time to mating effort over parental investment (Trivers, 1972; 2017). This became Parental Investment Theory and has been enormously influential in evolutionary psychology, informing both Sexual Strategies Theory and Life History Theory. Human males face a greater trade-off than human females when allocating time to mating effort (seeking sexual partners) or parental investment (commitment to one partner, among other things). This explains many sex differences in sexual behavior, such as the large sex differences we see in sociosexuality and mate preferences (Buss, 1989; Lippa, 2009).
Manosphere and red pill caricatures of “alphas” and “betas” map neatly onto low-investment and high-investment males, with regular reference to the so-called “beta provider.” However, “alpha” and “beta” are terms drawn from observations of animal status hierarchies. We should therefore expect to see low parental investment associated with high status (and high mate value) and high parental investment associated with low status (and low mate value). Do individual differences in parental investment really reflect this?
The answer is generally “no” on both levels of individual differences and demography.
Apicella & Marlowe (2007) found no direct relationship between mate value in men and parental investment. However, mating effort was associated with decreased parental investment and mediated the relationship between mate value and investment. This is consistent with life history theory; fast life history strategists have increased mating effort and lower parental investment.
Chang et al. (2017) found that traits indicative of “good genes” (masculinity, physical attractiveness, sexiness, etc.) were correlated at .5 with traits indicative of “good providers” (capable, high social status, high education, etc.). Rather than our folk psychology theory of traits — where smart people are unattractive, jocks are stupid, and sexy men are deadbeat losers — good behavioral and physical traits are positively (albeit often weakly) correlated. As explained in more recent, large-scale research, this is produced by assortative mating: high-value individuals select for both physical and behavioral high-value traits, producing a correlation between the two in offspring (Conroy-Beam 2018; Conroy-Beam et al., 2019). Further, life history strategy is also directly correlated with both physical attractiveness and mate value (Dillon et al., 2013). Slow life history strategists, or individuals who engage in high parental investment, are more physically attractive and have higher mate value on average.
Life history strategies are heritable (Figueredo et al., 2004) and also shaped by the developmental environment — in particular the age of sexual debut (Del Giudice, 2024). Therefore, we can also look at parental investment and offspring development. Individuals who have higher parental investment also have offspring with higher mate quality (Antfolk & Sjölund, 2018). In other words, individuals of high mate value are more likely to be long-term (or slow life history) strategists and are more likely to pass on traits that predispose offspring to parental investment (Kaptijn et al., 2010). High parental investment is associated with higher mate value in subsequent generations.
Buss & Shackelford (2008) also found that women of higher mate value (more physically attractive women) demand parental investment more than women of lower mate value. Attractive women are more likely to pick men who are committed and invested. They will share and pass on their “good genes” with the investors. In human beings, this will almost always be the primary partner, or the long-term relationship partner. Rates of extra-pair paternity (cuckoldry) are consistently between 1-3% (for a review of the literature, see Larmuseau et al. 2016). This is a powerful fact about human mating: the men that women are picking to pass their genes on with are the men they are selecting predominantly as long-term, not short-term, romantic partners.
Pair bonding and long-term, serially monogamous unions evolved long ago in the human line, with high estimates as late as Australopithecus afarensis 3 million years ago having been monogamous (Schacht & Kramer, 2019; Reno et al., 2003). Selection pressures early on may have acted against multiple mating and in favor of paternal investment with a single partner (Schacht & Bell, 2016). In the distant past (millions of years ago) it may have been the case that “lower value” ancestors of male humans used provisioning to secure mates rather than contest competitions, but they shaped human evolution in such a way that “higher value” male ancestors were induced to engage in provisioning as well (Gavrilets, 2012). In modern populations we should not necessarily expect a negative relationship between mate value and parental investment or provisioning, as male human ancestors of high and low value have been forced to compete on the dimensions of investment and commitment for millions of years. As described above, commitment and investment are simply desirable traits across the board (even for short-term mate selection) and, as such, we should expect them to correlate positively with many other desirable traits (including physically desirable traits).
Life History Strategies and Status
Buss et al. (2020) conducted a large study of high and low status traits across 14 nations. Here is what they found:
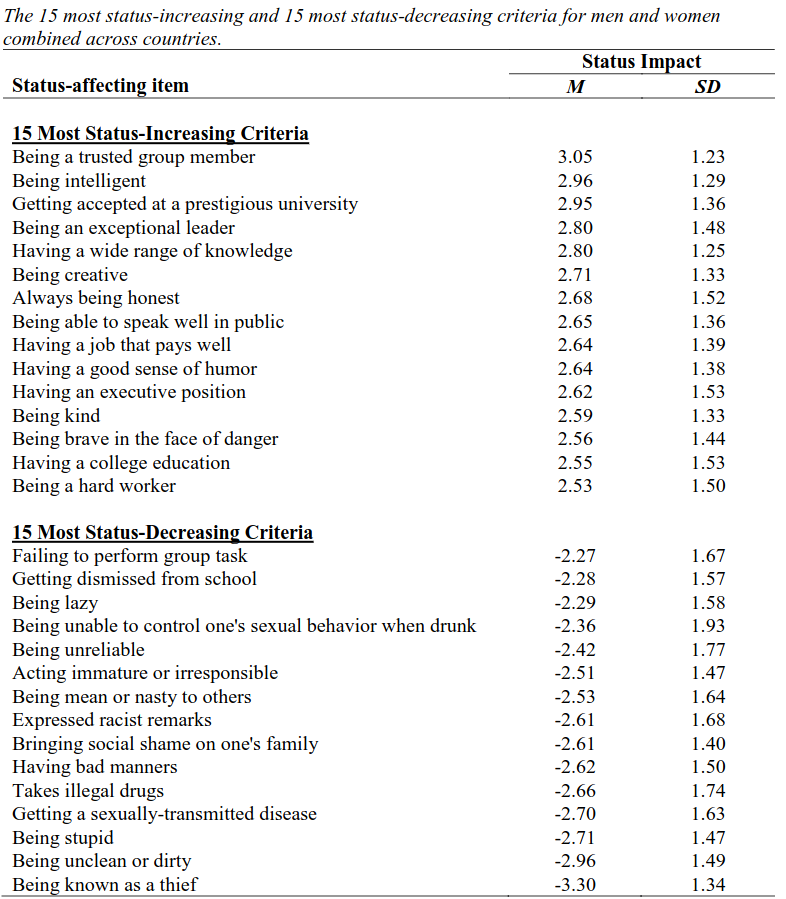
These align fairly well with fast and slow life history strategies, with the traits that more closely resemble slow life history strategies predicting higher status. Being an honest, trusted group member is generally cooperative and non-exploitative (Del Giudice, 2024), higher education is associated with a later timing at first birth and higher paternal investment (Hill & Kaplan, 1999; Bulled & Sosis, 2010; Waynforth, 2012), and substance abuse is associated with a fast life history (Hill & Chow, 2002). Antisociality broadly is associated with fast life histories (Gladden et al., 2009; Tielbeek, et al, 2018; Del Giudice, 2024) and behaviors or traits that confer high and low status essentially describe prosocial and antisocial traits respectively.
Poverty is associated with a present-orientation (fast) rather than a future-orientation (slow), as well as risky-behavior (Griskevicius et al., 2011). Parental investment and economic support predicts slow life history behaviors in adult offspring and higher status (Lukaszewski, 2015).
Life History, Development, and Romantic Relationship Outcomes
Given what you now know about Life History Theory we can conclude with an image from Jeffry Simpson illustrating associations between fast and slow life history strategies (2018) (image below). This is fitting given where this article began with the dual mating strategy hypothesis and ovulatory shift hypothesis, because Jeffry Simpson was the evolutionary psychologist alongside Steven Gangestad who came up with the strategic pluralism theory itself (2000).
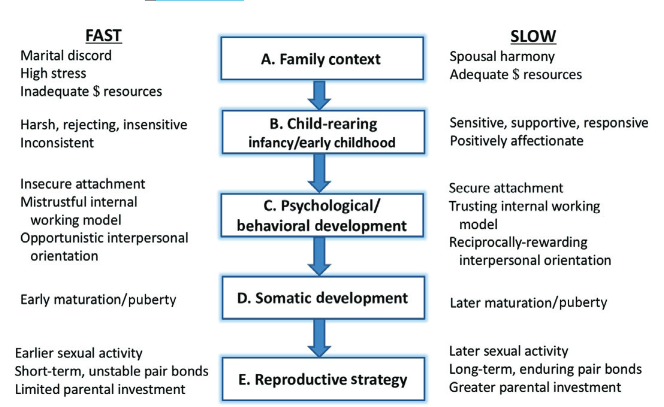
I won’t comment much on this, but ask yourself which of the two developmental trajectories and outcomes appears low or high value to you: a harmonious and resource-rich developmental environment that produces offspring of high somatic quality and enduring pair bonds, or its opposite?
Discussion
We began with a quick glance at what evolutionary psychologists think of manosphere takes on evolutionary psychology, using the dual mating strategy and ovulatory shift hypotheses to illustrate further how the manosphere misrepresents or misunderstands research in evolutionary psychology. From that we segued into a deeper dive on a common manosphere talking point: that men who form long-term pair bonds are “betas.”
Contrary to the claim that such men are “betas” we see that slow life history strategies and the formation of long-term pair bonds with high parental investment are associated with high mate value and high status.
Although I have pointed out many ways that popular red pill podcaster Rollo Tomassi gets evolutionary psychology wrong, here is something he has said that I like: high-value men are the men that other men want to be and the men that women desire.
This is actually a good take. Unfortunately, he and much of the manosphere gets it quite wrong when he associates antisocial behavior, a short-term mating orientation, and a fast life history strategy with high mate value.
Recently, Rollo Tomassi and denizens of the red pill subculture were sharing the story of Tavon White as an example of an “alpha.” You can read about him here. In summary, Tavon was a gang leader who impregnated four female guards, who he paid and recruited from the streets, while running an entire correctional facility from the inside.
Around the same time, the same influencers and denizens of the red pill were circulating the divorce story of Brazilian football player Ricardo Kaká. This was, for many, clear evidence that Kaká was a “beta.”
Kaká is a 6’1 professional athlete — one of Brazil’s most famous football players — and quite conventionally handsome. He maintained a monogamous relationship with his conventionally attractive wife and raised three children. Tavon White is not handsome, nor are the guards he impregnated conventionally attractive, and he resides within one of the lowest status environments in Western society: a prison.
Who do you think men would rather be and who do you think women would rather have sex with: Tavon White or Ricardo Kaká?
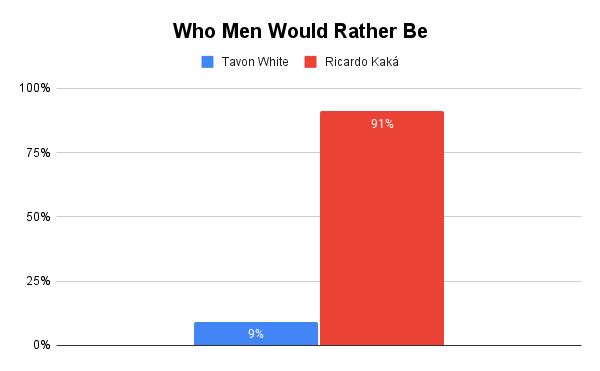
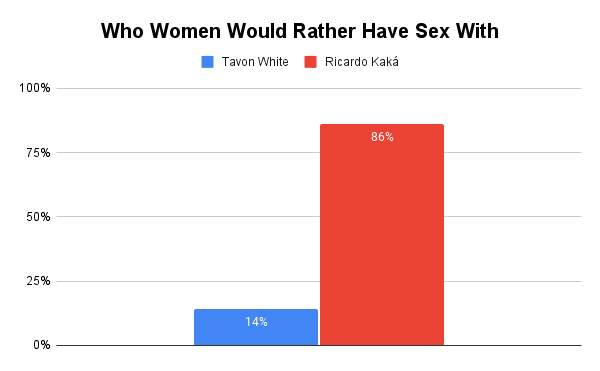
Unsurprisingly, our slow life history strategist beat our fast life history strategist.
I have written about this in the past (see: What the manosphere gets wrong about alphas and betas), but the caricature of the “alpha” as presented by the red pill often deviates from evolutionary perspectives on what high mate value really is.
Examples of men presented as “alphas” are often just men who pursue short-term mating strategies, who are high in antisocial traits, and/or who tend toward fast life history strategies. Actual markers of mate value, such as physical attractiveness or social status, are often discarded in lieu of merely having a “bad boy” vibe.
Why has Ricardo Kaká had fewer sexual partners than Travon White — is it because he has lower mate value? Kaká is an Evangelical Christian who married his high-school girlfriend and remained in a committed marriage for ten years. Travon White paid women to become prison guards so he could have sex with them. Individual differences in mating strategies produce high “body counts,” but as we have seen mate value is a poor predictor of which mating strategy men will enact. It was never the case that all men would pursue short-term strategies or enact fast life histories if they could.
Fast and slow life histories are produced by interactions between genes and the environment, with slow life history strategists having been dealt what is in many ways a better hand by mother nature. Individuals don’t simply choose fast life history strategies because they are more desirable and they don’t get forced into slow life history strategies because they are unattractive. In many ways, fast life history strategies are compensatory strategies for lower desirability. Fast life history strategists make compromises to the detriment of their own somatic effort and development, as well as compromises in choices of lower value mates, in order to attain mating opportunities.
References
Antfolk, J., & Sjölund, A. (2018). High parental investment in childhood is associated with increased mate value in adulthood. Personality and Individual Differences, 127, 144-150.
Apicella, C. L., & Marlowe, F. W. (2007). Men’s reproductive investment decisions: Mating, parenting, and self-perceived mate value. Human Nature, 18, 22-34.
Apostolou, M., & Michaelidou, E. (2024). Why people face difficulties in attracting mates: An investigation of 17 probable predictors of involuntary singlehood. Personality and Individual Differences, 216, 112422.
Arnocky, S. (2020). Mate-value moderates the relationship between intrasexual competitiveness and successful mate poaching. Evolutionary Psychological Science, 6(4), 346-353.
Arnocky, S., Desrochers, J., Rotella, A., Albert, G., Hodges-Simeon, C., Locke, A., … & Kelly, B. (2021). Men’s mate value correlates with a less restricted sociosexual orientation: a meta-analysis. Archives of Sexual Behavior, 50, 3663-3673.
Bachaud, L., & Johns, S. E. (2023). The use and misuse of evolutionary psychology in online manosphere communities: The case of female mating strategies. Evolutionary Human Sciences, 5, e28.
Boyce, V., Mathur, M., & Frank, M. C. (2023). Eleven years of student replication projects provide evidence on the correlates of replicability in psychology. Royal Society Open Science, 10(11), 231240.
Bulled, N. L., & Sosis, R. (2010). Examining the relationship between life expectancy, reproduction, and educational attainment: A cross-country analysis. Human Nature, 21, 269-289.
Buss, D. M. (1989). Sex differences in human mate preferences: Evolutionary hypotheses tested in 37 cultures. Behavioral and brain sciences, 12(1), 1-14.
Buss, D. M., & Shackelford, T. K. (1997). Susceptibility to infidelity in the first year of marriage. Journal of research in personality, 31(2), 193-221.
Buss, D. M., & Greiling, H. (1999). Adaptive individual differences. Journal of Personality, 67(2), 209-243.
Buss, D. M., & Shackelford, T. K. (2008). Attractive women want it all: Good genes, economic investment, parenting proclivities, and emotional commitment. Evolutionary Psychology, 6(1), 147470490800600116.
Buss, D. M., Durkee, P. K., Shackelford, T. K., Bowdle, B. F., Schmitt, D. P., Brase, G. L., … & Trofimova, I. (2020). Human status criteria: Sex differences and similarities across 14 nations. Journal of Personality and Social Psychology, 119(5), 979.
Buss, D. M. (2023). 1 The Sexual Selection of Human Mating Strategies: Mate Preferences and Competition Tactics. The Oxford Handbook of Evolutionary Psychology and Romantic Relationships, 15.
Buunk, A. P., & Massar, K. (2021). Intimate partner violence in Nicaragua: The role of possessive jealousy, intrasexual competitiveness, life history, mate value, and stress. Journal of interpersonal violence, 36(15-16), NP8101-NP8123.
Chang, L., Lu, H. J., & Zhu, X. Q. (2017). Good genes, good providers, and good fathers and mothers: The withholding of parental investment by married couples. Evolutionary Behavioral Sciences, 11(2), 199.
Conroy-Beam, D. (2018). Euclidean mate value and power of choice on the mating market. Personality and Social Psychology Bulletin, 44(2), 252-264.
Conroy-Beam, D., Roney, J. R., Lukaszewski, A. W., Buss, D. M., Asao, K., Sorokowska, A., … & Zupančič, M. (2019). Assortative mating and the evolution of desirability covariation. Evolution and Human Behavior, 40(5), 479-491.
Conroy-Beam, D., Buss, D. M., Asao, K., Sorokowska, A., Sorokowski, P., Aavik, T., … & Zupančič, M. (2019). Contrasting computational models of mate preference integration across 45 countries. Scientific reports, 9(1), 16885.
Davies, A. P., & Shackelford, T. K. (2017). Don’t you wish your partner was hot like me?: The effectiveness of mate poaching across relationship types considering the relative mate-values of the poacher and the partner of the poached. Personality and Individual Differences, 106, 32-35.
Del Giudice, M. A Turning Point for the Life History Approach to Individual Differences.
Dillon, H. M., Adair, L. E., Wang, Z., & Johnson, Z. (2013). Slow and steady wins the race: Life history, mate value, and mate settling. Personality and Individual Differences, 55(5), 612-618.
Dinh, T., & Gangestad, S. W. (2024). Mating fast and slow? Sociosexual orientations are not reflective of life history trajectories. Evolution and Human Behavior, 45(1), 27-40.
Dinh, T., Haselton, M. G., & Gangestad, S. W. (2022). “Fast” women? The effects of childhood environments on women’s developmental timing, mating strategies, and reproductive outcomes. Evolution and Human Behavior, 43(2), 133-146.
Ellis, B. J., Figueredo, A. J., Brumbach, B. H., & Schlomer, G. L. (2009). Fundamental dimensions of environmental risk: The impact of harsh versus unpredictable environments on the evolution and development of life history strategies. Human nature, 20, 204-268.
Figueredo, A. J., Vasquez, G., Brumbach, B. H., & Schneider, S. M. (2004). The heritability of life history strategy: The k‐factor, covitality, and personality. Social biology, 51(3-4), 121-143.
Figueredo, A. J., Vásquez, G., Brumbach, B. H., Schneider, S. M., Sefcek, J. A., Tal, I. R., … & Jacobs, W. J. (2006). Consilience and life history theory: From genes to brain to reproductive strategy. Developmental Review, 26(2), 243-275.
Gildersleeve, K., Haselton, M. G., & Fales, M. R. (2014). Do women’s mate preferences change across the ovulatory cycle? A meta-analytic review. Psychological bulletin, 140(5), 1205.
Hill, E. M., & Chow, K. (2002). Life‐history theory and risky drinking. Addiction, 97(4), 401-413.
Hill, K., & Kaplan, H. (1999). Life history traits in humans: Theory and empirical studies. Annual review of anthropology, 28(1), 397-430.
Jonason, P. K. (2008). A mediation hypothesis to account for the sex difference in reported number of sexual partners: An intrasexual competition approach. International Journal of Sexual Health, 19(4), 41-49.
Jonason, P. K., & Fisher, T. D. (2009). The power of prestige: Why young men report having more sex partners than young women. Sex Roles, 60, 151-159.
Jonason, P. K., & March, E. (2023). The Three Cs of Psychological Mate Preferences: The Psychological Traits People Want in Their Romantic and Sexual Partners. The Oxford Handbook of Evolutionary Psychology and Romantic Relationships, 74.
Gangestad, S. W. (1993). Sexual selection and physical attractiveness: Implications for mating dynamics. Human Nature, 4, 205-235.
Gangestad, S. W., & Simpson, J. A. (2000). The evolution of human mating: Trade-offs and strategic pluralism. Behavioral and brain sciences, 23(4), 573-587.
Gavrilets, S. (2012). Human origins and the transition from promiscuity to pair-bonding. Proceedings of the National Academy of Sciences, 109(25), 9923-9928.
Gladden, P. R., Figueredo, A. J., & Jacobs, W. J. (2009). Life history strategy, psychopathic attitudes, personality, and general intelligence. Personality and Individual Differences, 46(3), 270-275.
Griskevicius, V., Tybur, J. M., Delton, A. W., & Robertson, T. E. (2011). The influence of mortality and socioeconomic status on risk and delayed rewards: a life history theory approach. Journal of personality and social psychology, 100(6), 1015.
Kaptijn, R., Thomese, F., van Tilburg, T. G., Liefbroer, A. C., & Deeg, D. J. (2010). Low fertility in contemporary humans and the mate value of their children: sex-specific effects on social status indicators. Evolution and Human Behavior, 31(1), 59-68.
Kennison, S. M., Byrd-Craven, J., & Hamilton, S. L. (2017). Individual differences in talking enjoyment: The roles of life history strategy and mate value. Cogent Psychology, 4(1), 1395310.
Larmuseau, M. H., Matthijs, K., & Wenseleers, T. (2016). Cuckolded fathers rare in human populations. Trends in ecology & evolution, 31(5), 327-329.
Li, N. P., & Choy, B. K. (2023). Partner evaluation and selection. In The Oxford handbook of evolutionary psychology and romantic relationships (pp. 94-126). Oxford University Press.
Lippa, R. A. (2009). Sex differences in sex drive, sociosexuality, and height across 53 nations: Testing evolutionary and social structural theories. Archives of sexual behavior, 38, 631-651.
Lukaszewski, A. W. (2015). Parental support during childhood predicts life history-related personality variation and social status in young adults. Evolutionary Psychological Science, 1, 131-140.
MacArthur, R. H., & Wilson, E. O. (2001). The theory of island biogeography (Vol. 1). Princeton university press.
McDowell, J., & Starratt, V. G. (2021). Positive correlation between high mate value and slow life history strategy is moderated by trait heritability. Biodemography and Social Biology, 66(1), 27-39.
Michal, A. L., & Shah, P. (2024). A Practical Significance Bias in Laypeople’s Evaluation of Scientific Findings. Psychological Science, 35(4), 315-327.
Michaelidou, M. A. E. Good looks, mate value, flirting capacity, and choosiness: Exploring additional pathways to singlehood.
Miner, E. J., Starratt, V. G., & Shackelford, T. K. (2009). It’s not all about her: Men’s mate value and mate retention. Personality and Individual Differences, 47(3), 214-218.
Murasko, J. E. (2020). Height, marriage, and partner characteristics for women in low-and middle-income countries. Economics & Human Biology, 38, 100876.
Nascimento, B. S., Adair, L., & Vione, K. (2024). Pathways to online infidelity: the roles of perceived online dating success, perceived availability of alternative partners, and mate value discrepancy. Current Psychology, 43(14), 12782-12793.
Nijboer, A. C. S., Sellitto, M., Ruitenberg, M. F. L., Kerkkänen, K. I. L., & Schomaker, J. (2024). Food-related exploration across the menstrual cycle. Appetite, 107261.
Reno, P. L., Meindl, R. S., McCollum, M. A., & Lovejoy, C. O. (2003). Sexual dimorphism in Australopithecus afarensis was similar to that of modern humans. Proceedings of the National Academy of Sciences, 100(16), 9404-9409.
Rushton, J. P. (2004). Placing intelligence into an evolutionary framework or how g fits into the r–K matrix of life-history traits including longevity. Intelligence, 32(4), 321-328.
Schacht, R., & Bell, A. V. (2016). The evolution of monogamy in response to partner scarcity. Scientific reports, 6(1), 32472.
Schacht, R., & Kramer, K. L. (2019). Are we monogamous? A review of the evolution of pair-bonding in humans and its contemporary variation cross-culturally. Frontiers in Ecology and Evolution, 7, 426706.
Schleifenbaum, L., Driebe, J. C., Gerlach, T. M., Penke, L., & Arslan, R. C. (2021). Women feel more attractive before ovulation: Evidence from a large-scale online diary study. Evolutionary Human Sciences, 3, e47.
Schmitt, D. P., & Buss, D. M. (2001). Human mate poaching: Tactics and temptations for infiltrating existing mateships. Journal of personality and Social Psychology, 80(6), 894.
Schmitt, D. P. (2004). Patterns and universals of mate poaching across 53 nations: the effects of sex, culture, and personality on romantically attracting another person’s partner. Journal of Personality and Social Psychology, 86(4), 560.
Sela, Y., Mogilski, J. K., Shackelford, T. K., Zeigler‐Hill, V., & Fink, B. (2017). Mate value discrepancy and mate retention behaviors of self and partner. Journal of Personality, 85(5), 730-740.
Shackelford, T. K., Schmitt, D. P., & Buss, D. M. (2005). Universal dimensions of human mate preferences. Personality and individual differences, 39(2), 447-458.
Simpson, J. A., & Gangestad, S. W. (1991). Individual differences in sociosexuality: evidence for convergent and discriminant validity. Journal of personality and social psychology, 60(6), 870.
Simpson, J. A. (2019). Bringing life history theory into relationship science. Personal Relationships, 26(1), 4-20.
Starratt, V. G., Weekes-Shackelford, V., & Shackelford, T. K. (2017). Mate value both positively and negatively predicts intentions to commit an infidelity. Personality and Individual Differences, 104, 18-22.
Steckel, R. H. (2009). Heights and human welfare: Recent developments and new directions. Explorations in Economic History, 46(1), 1-23.
Stephen, I. D., & Luoto, S. (2021). Physical cues of partner quality. The Oxford handbook of evolutionary psychology and romantic relationships.
Thomas, A. G., Armstrong, S. L., Stewart-Williams, S., & Jones, B. C. (2021). Current fertility status does not predict sociosexual attitudes and desires in normally ovulating women. Evolutionary Psychology, 19(1), 1474704920976318.
Tielbeek, J. J., Barnes, J. C., Popma, A., Polderman, T. J., Lee, J. J., Perry, J. R., … & Boutwell, B. B. (2018). Exploring the genetic correlations of antisocial behaviour and life history traits. BJPsych open, 4(6), 467-470.
Trivers, R. L. (2017). Parental investment and sexual selection. In Sexual selection and the descent of man (pp. 136-179). Routledge.
Walter, K. V., Conroy-Beam, D., Buss, D. M., Asao, K., Sorokowska, A., Sorokowski, P., … & Zupančič, M. (2020). Sex differences in mate preferences across 45 countries: A large-scale replication. Psychological Science, 31(4), 408-423.
Waynforth, D. (2012). Life-history theory, chronic childhood illness and the timing of first reproduction in a British birth cohort. Proceedings of the Royal Society B: Biological Sciences, 279(1740), 2998-3002.
Wood, W., Kressel, L., Joshi, P. D., & Louie, B. (2014). Meta-analysis of menstrual cycle effects on women’s mate preferences. Emotion Review, 6(3), 229-249.
Zhuang, J. Y., & Wang, J. X. (2014). Women ornament themselves for intrasexual competition near ovulation, but for intersexual attraction in luteal phase. PloS one, 9(9), e106407.
3 comments
That last paragraph is funny because it shows that redpillers are still right to pursue and support short-term/fast-life strategies even though they’re wrong about the “why”. Most redpillers are unattractive and therefore SHOULD pursue fast-life strategies.
I have an off-topic question when it comes to the white card dispute with genetics, tell me, what do you think about wild babies?
of course human wild babies https://en.wikipedia.org/wiki/Victor_of_Aveyron